Gap costs
The alignment algorithm has three parameters
concerning gap costs: Gap open cost, Gap extension cost and End gap
cost. The precision of these parameters is to one place of decimal.
- Gap open cost. The price for introducing gaps in an alignment.
- Gap extension cost. The price for every extension past the initial gap.
If you expect a lot of small gaps in your alignment, the Gap open
cost should equal the Gap extension cost. On the other hand, if you
expect few but large gaps, the Gap open cost should be set
significantly higher than the Gap extension cost.
However, for most alignments it is a good idea to make the Gap open
cost quite a bit higher than the Gap extension cost. The default
values are 10.0 and 1.0 for the two parameters, respectively.
- End gap cost. The price of gaps at the beginning or the end of the alignment.
One of the advantages of the CLC Genomics Workbench alignment method is that it
provides flexibility in the treatment of gaps at the ends of the
sequences. There are three possibilities:
- Free end gaps. Any number of gaps can be inserted in the ends of
the sequences without any cost.
- Cheap end gaps. All end gaps are treated as gap extensions and
any gaps past 10 are free.
- End gaps as any other. Gaps at the ends of sequences are treated
like gaps in any other place in the sequences.
When aligning a long sequence with a short partial sequence, it is
ideal to use free end gaps, since this will be the best
approximation to the situation. The many gaps inserted at the ends
are not due to evolutionary events, but rather to partial data.
Many homologous proteins have quite different ends, often with large
insertions or deletions. This confuses alignment algorithms, but
using the Cheap end gaps option, large gaps will generally be
tolerated at the sequence ends, improving the overall alignment.
This is the default setting of the algorithm.
Finally, treating end gaps like any other gaps is the best option
when you know that there are no biologically distinct effects at the
ends of the sequences.
Figures 20.3 and 20.4 illustrate the
differences between the different gap scores at the sequence ends.

Figure 20.3: The first 50 positions of two different alignments of seven calpastatin sequences. The top alignment is made with cheap end gaps, while the bottom alignment is made with end gaps having the same price as any other gaps. In this case it seems that the latter scoring scheme gives the best result.
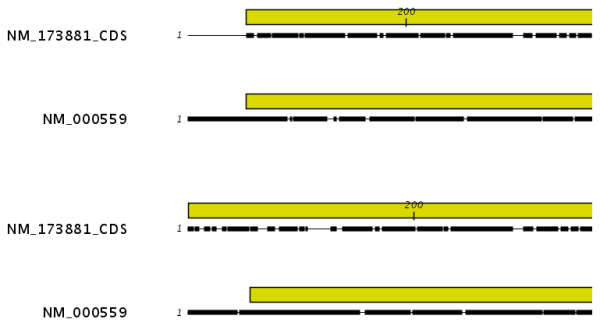
Figure 20.4: The alignment of the coding sequence of bovine myoglobin with the full mRNA of human gamma globin. The top alignment is made with free end gaps, while the bottom alignment is made with end gaps treated as any other. The yellow annotation is the coding sequence in both sequences. It is evident that free end gaps are ideal in this situation as the start codons are aligned correctly in the top alignment. Treating end gaps as any other gaps in the case of aligning distant homologs where one sequence is partial leads to a spreading out of the short sequence as in the bottom alignment.